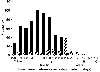What's so special about drylands?
![]() No.
59, August 2007
No.
59, August 2007
What's so special about drylands?
by Mary Price and Nick Waser
|
"Understanding patterns of variation and unpredictability, and how individual species... cope with these key characteristics of deserts, leads to an understanding of entire desert ecosystems." |
|
|
|
Deserts defined(Back to top) Deserts are one of the Earth's major ecosystems, occupying about 30% of the land surface and occurring around 30 degrees latitude on all continents. Technical definitions of deserts are couched in terms of averages. They are drylands with the emphasis on dry, regions that receive less than a certain cutoff amount of annual precipitation–250 mm (~10 inches) according to McGinnies et al.'s (1970) classification scheme. But this definition doesn't exclude places like Alaskan tundra, which meets the precipitation cutoff but isn't considered a desert because water accumulates in permafrost. A better definition takes water loss into account: a desert is a dryland where potential evapotranspiration exceeds annual precipitation. This, too, is a definition in terms of average conditions. |
|
|
Charleville's houses on stilts tell us that deserts aren't always dry. What definitions based on averages miss is variation and unpredictability. Precipitation in deserts comes in discrete pulses–-"events" of variable size that often are brief, lasting minutes or hours rather than days (Figure 4), and that are separated by droughts of unpredictable length. The entire annual input may come in a single rain event, or in 50 events, and years, or months, may pass between successive events. Water can be plentiful in the arid center of Australia--but only at some times, and in some places. Of course, if this were not so there would be very little desert life! So deserts might best be defined as "water-controlled ecosystems with infrequent, discrete, and largely unpredictable water inputs" (Noy-Meir 1973). Exploiting moisture, enduring drought(Back to top) Other desert organisms are truly desert-adapted, having evolved strategies for coping with sparse and unpredictable moisture by avoiding the driest periods or by enduring them. Although we and others have tended to characterize these adaptations as either/or--that is, strategies that avoid drought vs. those that endure drought--in reality, avoidance and endurance strategies grade into one another and probably are best thought of as end points on a continuum. The purest "drought avoiders" are species with clever mechanisms for being biologically active only when moisture is available. Some, like the ephemeral plants responsible for the spectacular wildflower displays in wet years in some deserts, persist during drought as dormant seeds in the soil. Their primary desert adaptations involve mechanisms for germinating quickly in response to cues that signal an upcoming period of favorable growth conditions, and for having some fraction of seeds remain dormant as a form of "bet-hedging" in case the growth period turns out to be too short to produce a new crop of seeds. Once they germinate, the ephemerals' strategy is to complete seed production during the brief moist window of opportunity; they lack special adaptations for living through drought as vegetative plants, and so they die young when drought returns. Some such species grow into large plants and flower profusely if moisture persists, whereas they can respond by flowering when only a few centimeters tall if conditions dry up soon after they germinate as seedlings (the Australian parakeelyas, Calandrinia species, are good examples). Similarly, the spadefoot toads of North American deserts (Scaphiopus species) and spadefoot frogs of Australian deserts (Notaden and Neobatrachus species) wrap themselves in waterproof cocoons and go dormant underground until it rains again. A different "avoider" strategy, which is only possible for more mobile organisms, is to move to more favorable places once drought sets in. Seasonal migrants such as birds and some insects do this in the deserts of North America, passing through them in moist seasons. But in deserts where moisture is less predictable seasonally, regular migration is replaced by more opportunistic regional movements-i.e., by nomadism. Many Australian birds and even some small mammals and reptiles follow moisture in space. Their geographic distributions include the entire arid center of the continent, but at any one time they are found only in places that have recently received precipitation. Similarly, desert peoples often are nomadic, moving with their stock or game to exploit rain-flushed vegetative growth and rain-filled waterholes. In turn, the purest "drought endurers" are species that stay put and remain physiologically active at some reasonably high level when drought sets in (the fact that dormant seeds or spadefoot toads and frogs are also metabolically active, albeit at a very low level, is an example of how endurer and avoider strategies grade into one another). They do so by having adaptations for taking up and storing water when it is available, and using it sparingly during drought. Physiological ecologists have uncovered a rich and wonderful diversity of ways in which this basic strategy unfolds among desert organisms. For example, perennial desert plants can minimize evaporative water loss from their leaves by being drought-deciduous; or by having small, pale leaves that reflect solar radiation and shed heat; or by taking up carbon dioxide for photosynthesis during the cool hours of the night. |
|
|
Desert animals have a similar bag of tricks. To take an example from our own North American deserts, the kangaroo rats (Dipodomys species), and their relatives the kangaroo mice (Microdipodops species) and pocket mice (Perognathus and Chaetopdipus species), obtain both water and food from the seeds that they collect in moist periods when plants reproduce, and that they then store (cache) in the soil (Figure 5). These small rodents can live solely on the "metabolic water" that is produced when they digest the seeds (carbohydrate, fat, and protein produce carbon dioxide, water, and energy as the end products of digestion). This feat is possible because kangaroo rats and their allies are superb water misers: they live in humid burrows, are active aboveground only at night when it is cool (so they do not have to use water to thermoregulate), and have noses and kidneys designed to retain water. Variable rainfall: Key to biodiversity patterns |
|
|
(Back to top) Second, no organismal strategy is effective at exploiting all sizes of precipitation event. For example, small rains that moisten just the soil surface can only be exploited by short-lived organisms that can respond quickly and go dormant again quickly, because moisture soon evaporates from shallow soil layers. Cues that trigger seed germination are highly species-specific, so that any particular pattern of moisture input will be available to only a subset of plant species. The deep moisture that comes from large events in turn can only be tapped by deep roots. Many succulent plants cannot use cold-season moisture. And large animals need rain events large enough to fill up large waterholes. Hence, when rainfall is variable, no one organism can use all the moisture and exclude all other species--some is left for use by others with different strategies. A corollary is that the particular mix of organismal strategies will vary among deserts that have different patterns of water input. Fog deserts such as the Vizcaíno of Baja California, Mexico, and the Namib of southwestern Africa, have an abundance of rootless lichens and micro-organisms that are rare elsewhere. Deserts with long and variable return times, such as the Simpson Desert of Australia, support a high proportion of ephemeral plants. Deserts compared(Back to top) |
|
|
We work on kangaroo rats and their pocket mouse relatives, members of the rodent family Heteromyidae, which are dominant nocturnal seed-eating animals in our hot deserts (Figure 7). We were interested in seeing the Australian equivalents of our heteromyid rodents-the hopping mice and false mice (Figure 8). What impressed us were the differences, rather than the similarities, between these so-called ecological equivalents. Although hopping mice resemble kangaroo rats in hopping like tiny kangaroos, and false mice resemble pocket mice, their behaviors are very different. Kangaroo rats and pocket mice are sedentary and solitary. They live in underground burrows that take a great deal of energy to build, and they have evolved cheek pouches and elaborate behaviors for gathering and storing seeds, for stealing seeds from other rodents, and for protecting their own seed caches from such thieves. Australian desert mice are similar only in that they eat seeds, and even then seeds seldom form more than two-thirds of the diet. Hopping mice live in burrows, to be sure, but the usual burrows are cheap temporary structures. Breeding burrows are deeper and more impressive, but are constructed only in times of plenty. These animals also lack specialized pouches for transporting seeds. They store seeds for immediate consumption only, are quite social, and are anything but sedentary. Our colleague Dr. Chris Dickman of the University of Sydney has documented that these animals regularly move several kilometers in a night-something unheard of for any self-respecting adult kangaroo rat! Another difference is that Australian desert rodents show "irruptive" population dynamics, in which there are no animals at all in a locality for long periods of time, and then suddenly populations reach huge densities for a short time before crashing back to zero. North American heteromyid populations, in contrast, fluctuate much less wildly. We concluded that the "heteromyid niche" simply doesn't exist in Australia. Why not? The Simpson Desert has approximately the same average annual rainfall as North American deserts. But the variance in rainfall is considerably greater, and the "return times" of significant seed-generating rain events at any single locality are longer. Furthermore, Australia is an old continent with weathered, nutrient-poor soils. It takes a larger water input to achieve plant productivity equivalent to that seen in nutrient-rich North American soils. Perhaps a single rodent in a single territory cannot expect to be able to harvest and manage enough seeds to last until the next pulse of rain and seed production in that location. Perhaps the only viable strategy for a rodent in arid Australia is to be nomadic. The exceptional variance and longer return times for precipitation events, soil infertility, and flat topography of arid Australia appear to be responsible for many of its unique ecological features. These include a general dominance by non-succulent perennial sclerophyllous plants and by termites and termite-eating lizards compared to groups that are abundant in other deserts, such as ephemeral plants, leaf-feeding insects, and mammals. Extended to human cultures, these same unique features of Australian deserts may explain why desert aboriginals were and are nomadic and rely on fire-with its attendant nutrient input-rather than on flood agriculture, in contrast to the sedentary O'odham peoples of the Sonoran Desert, who cultivate crops in summer flood catchments. And of course, these features explain the vortex guns and stilts of towns in the Queensland Channel Country. The future of variation(Back to top) The climate of Planet Earth is changing, and it now is apparent even to non-scientists that human activities (in particular our release of greenhouse gases, mostly from burning of fossil fuels) are responsible. As the climate warms, the human population of drylands, and the myriad other organisms on which they depend, will need to deal with increased variation and unpredictability (Reynolds et al. 2007). The best scientific information now available suggests that variation and unpredictability in water inputs to deserts will grow over the foreseeable future (Meehl et al. 2007). What can we learn from what we know at this point about deserts, their differences around the planet, and the strategies of plants, animals and humans that have adapted to them and to their inherent variability? The answers will provide keys as to the sustainability of human occupation of the deserts of the next century. Acknowledgments(Back to top) References(Back to top) Meehl, G. A., T. F. Stocker, W. D. Collins, et al. 2007. Global climate projections. In Climate Change 2007, the Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, eds. S.Solomon, D. Qin, M. Manning, et al., 747-845. Cambridge, UK: Cambridge University Press. Noy-Meir, I. 1973. Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics 4:25-51. Reynolds, J. F., D. M. Stafford Smith, E. F. Lambin, et al. 2007. Global desertification: building a science for dryland development. Science 316:847-851. Stafford Smith, D. M. and S. R. Morton. 1990. A framework for the ecology of arid Australia. Journal of Arid Environments 18:255-278. |
![]()
(Back to top)
Mary Price (mary.price@ucr.edu) and
Nick Waser (nickolas.waser@ucr.edu)
are adjunct professors in the School of Natural Resources, the University of
Arizona, and biology professors emeritus at the University of California, Riverside.
(Back to top)
Living in a variable climate
http://www.environment.gov.au/soe/2006/publications/integrative/climate/index.html
Index page for a multi-part integrative commentary on Australia's climate,
commissioned by the 2006 Australian State of the Environment Committee, 2006
Impact of climate change and land use in the Southwestern United States: Papers
on climatic variability
http://geochange.er.usgs.gov/sw/ataglance.html#climvar
Index page linking to
papers on climate variability from a 1997 workshop sponsored by the U.S. Department
of the Interior and the U.S. Geological Survey.
About the Arid Lands Newsletter