![[Previous]](prevpage.gif)
![[Next]](d.nextpage.gif)
Session leader: Dave Bellamy.


Hexapod size varies more than in any other class of animals, with length variations covering four orders of magnitude, and mass variations of seven orders of magnitude for extant hexapods. It is important to note that the "range" discussed in this section has nothing to do with "limits" in body size. Just because we know of no hexapod to ever have been larger than 750 mm does not necessarily mean that hexapods cannot be larger. The same holds true for the smaller hexapods. Possible limiting factors will be discussed in the following sections and during the class discussion.
Some of the smallest hexapods are members of springtails (Neelidae) (0.27 mm), thrips (0.5 mm), minute bog beetles (Sphaeriidae) (0.5 mm), feather-winged beetles (Ptiliidae) (0.25 mm), and many Chalcidoidea (Hymenoptera) that include the smallest members, fairyflies (Mymaridae) (<< 0.5 mm) and Trichogrammatidae (0.18 mm). The Trichogrammatidae are parasites of thrip eggs and may have a mass less than one microgram!
Some of the largest insects only flourished during the Carboniferous period (350 mya). This was a period dominated by many large dragonfly-like insects, including Meganeuropsis permiana Carpenter, within the extinct order Meganisoptera. This monster had a wingspan between 710 mm and 750 mm (or about 28 inches), making it the largest known insect ever to have existed. Click here to see photos of other fossil dragonflies.
Some of the largest extant hexapods include dragonflies (Aeshnidae)(135 mm), Phasmida (310 mm), the Chinese mantid (Montodea)(>100 mm), some tropical species of queen termites (Isoptera)(110 mm), giant water bugs (Belostomatidae)(>100 mm), a tropical plant hopper (Fulgoridae)(150 mm wingspread), dobsonflies (Corydalidae)(130 mm wingspread), and a number of beetles (Dynastinae, Cetoniinae, Cerambycidae, Buprestidae, etc.)(>100 mm). Click here to see photos of some of the beetles mentioned, including the Goliath beetle with a mass of 100 grams!
Now that the range has been examined, we can speculate as to whether this range is actually the limit for the hexapod body plan. Is this limit a physical one or an environmental one? Both categorical limitations could exist, but surely one is encountered before the other. Which one, then, has more of an impact on hexapods? What factors might actually account for the range in size and mass that we see? Have these factors always existed, or are they newly imposed? Are these factors universal within the animal kingdom, or are they unique to the insect world? Because of the scarcity of definitive literature on this topic, the answers to these questions currently remain speculative. Here are some ideas that have been put forth within the literature (and some through discussions).
What might be the physical factors that regulate maximum body size in insects? The two prevailing ideas include the limitations of the tracheal respiratory system and limitations of exoskeleton growth and strength.
Respiration in insects is dependent upon two distinct phases. First, oxygen must diffuse from the environment to the furthest reaches of the tracheal system. This phase is referred to as air-tube diffusion and is due to a difference in partial pressures between the internal and external environment. Once a differential in partial pressures is established (a drop as little as 2% of an atmosphere is sufficient), oxygen diffuses throughout the tracheal system (Chapman, 1969). Secondly, oxygen must then diffuse from the tracheoles into the tissues and ultimately to the mitochondria. This second step is know as tissue diffusion and is by far the slower of the two phases, since oxygen diffuses some 100,000 times faster in air than it does in the tissues (Chapman, 1969). The rate of diffusion through tissues, therefore, limits the distance any tissue can be from a tracheole, and consequently, the size of the of the tissue (e.g. muscle fibers, etc.). Diffusion can provide enough oxygen for the demands of small insects, but alone, it cannot meet the demands of highly active tissue in larger insects (Chapman, 1969). Larger insects, therefore, rely on ventilation (convection produced by changes in the tracheal system volume) to supplement their need. But even this is not enough to provide needed oxygen during flight for large insects. It appears as though large insects have resorted to using every technique available to them to harvest needed oxygen, including the contraction and expansion of the thorax, and using changes in the muscle volume to compress intramuscular air sacs (Chapman, 1969). So, indeed, it looks as though the insects are approaching the physiological limit of the tracheal system. For additional information on the tracheal system and insect respiration, click here for a lecture by John Pinto of UC Riverside.
It may also be that having a cuticle that must be molted to grow has inherent ecological and physical limitations. Hutchinson, et al (1997) state that optimum size increases in Arthropods are a function of the scaling of feeding rate with size, the efficiency of molting and the optimum size increase at the preceding molt. Hutchinson, et al, believe that there is an optimum duration of each instar and an optimum number of instars. This holds true for especially for insects, whose feeding rate depends on the size of structures (i.e., mandibles) that increase discontinually (only at molts). They hold that while the abdomen can expand to hold reserves while feeding, the rate at which many insects can ingest food is dependent upon some inextensible structure(s). Furthermore, the size of these structures depend on how much they grew during the last molt, which depended on how much reserves were accumulated before that molt, which in turn depended on the feeding rate. If in fact, there is a strategy for optimizing growth in insects, there must be some selective advantage to growing as big as possible, efficiently. Do insects express their ecologically maximum growth rate? ** Click here for a side trip for those interested in some amazing slides showing the metamorphosis of one of the largest insects in the world. **
It is much easier to speculate on factors that might limit maximum hexapod size than for minimum body size. Very little information exists, in a written form, that examines the "smallness" of hexapods. However, most entomologists are willing to offer hypothetical views of possible limiting factors. Here are the most entertaining of those ideas.
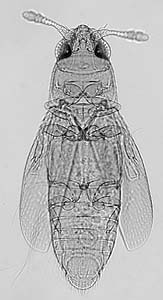 The
most plausible idea is that the size of the egg during its development
places a lower limit on female size. Insect eggs are relatively large because
they contain a large amount of yolk needed for the developing larvae. Some
parasitic insects have no need for yolk since the larva develop within
the nutrient rich fluid of their host, and can therefore, produce smaller
eggs. The eggs of some fairyflies (Mymaridae) can be as small as 0.06 mm
in length. Small eggs, however, are not confined to parasitic insects.
The feather-wing beetles of the genus Nanosella (shown left) are
so small (.25 mm) that females can only produce one egg at a time within
the abdominal region, since egg size is generally half the body length
(Ptiliidae photo and information from Tree
of Life and Gene Hall). Perhaps egg viability, known to be dependent
upon the female's dietary history, has set the lower boundary for insect
size. Smaller eggs, simply aren't as likely to survive.
The
most plausible idea is that the size of the egg during its development
places a lower limit on female size. Insect eggs are relatively large because
they contain a large amount of yolk needed for the developing larvae. Some
parasitic insects have no need for yolk since the larva develop within
the nutrient rich fluid of their host, and can therefore, produce smaller
eggs. The eggs of some fairyflies (Mymaridae) can be as small as 0.06 mm
in length. Small eggs, however, are not confined to parasitic insects.
The feather-wing beetles of the genus Nanosella (shown left) are
so small (.25 mm) that females can only produce one egg at a time within
the abdominal region, since egg size is generally half the body length
(Ptiliidae photo and information from Tree
of Life and Gene Hall). Perhaps egg viability, known to be dependent
upon the female's dietary history, has set the lower boundary for insect
size. Smaller eggs, simply aren't as likely to survive.
Another possible factor returns to the structure of the feeding mechanisms. Simply put, particular feeding styles (xylem feeding, eating pollen grains, etc.) require mouthparts strong enough to get at or manipulate that food source. Furthermore, the head of the insect must house the muscles necessary to operate these mouthparts. Novotny and Wilson (1997) use this argument, rather effectively, to explain the lower size limits for xylem feeding insects. Perhaps this is why many small insects are not mandibulate (For a nice mosquito picture and a diagram of its internal mouthparts click here . There is also a great site discussing whitefly mouthparts with pictures -- click here ). While the idea of mouthparts and feeding habit placing a lower boundary on size sounds likely for particular groups of insects, there is no consistent pattern among the hexapods.
The theory that animal body sizes change over time has been examined extensively (for vertebrates!). The apparent tendency for body size to increase over time in many diversifying taxonomic groups has been referred to as Cope's Rule (Cope, 1887). Explanations for this trend have usually centered around specific advantages of large individuals, such as ability to tolerate short-term variation in the physical environment, capacity to extract energy and nutrients from a wider variety of poorer quality food, and ability to avoid many kinds of predators. Cope's Rule has usually been used in the discussion of birds and mammals. What trends do we see in the hexapod world? Does Cope's Rule hold for insects as well? If not, why not?
Many entomologists might say that hexapods have been getting smaller. Perhaps this is due to the brief presence of the giant Protodonates during the Carboniferous period. It would be easy to draw a conclusion from the fossil evidence that dragonflies have become smaller over time. Indeed, many of the largest insects known to exist flourished during the Carboniferous only to become extinct. However, the presence of giant insects does not exclude the presence of small insects. Actually, the oldest known fossil hexapod is a springtail (Rhyniella praecursor Hirst & Maulik) from the lower Devonian. Scourfield (1940) judged from the length of the head, that the specimen appears to have been from 0.3 to 0.4 mm, a size that corresponds closely with those Collembolans living today. In fact many fossil records show that many hexapods have remained consistent in size throughout their existence. Not only did many Carboniferous giants become extinct, but so did their orders. Therefore, one cannot argue that these orders have become smaller over time, they simply were unsuccessful as insects and died out, perhaps allowing for future increases in hexapod diversity.
All this speculation on factors that limit size may be important when addressing the issue of insect diversity. Hexapods may be the most diverse and numerous -- both in terms of species richness and individual population -- life form on Earth. How have they managed to succeed so well? It has been suggested in many introductory biology textbooks that the ability to diversify depends on the homogeneity of the environment. The larger an animal is, the more homogenous their environment appears in terms of available niches. Smaller animals, on the other hand, live in environments rich with possible niches. However, this does not mean it's better to get smaller. As we have seen, there are many influential limiting factors present in the environment.
Siemann et al (1996), examined species diversity, abundance and body size relationships in an grassland insect community and found that the intermediate body sizes for each sampled order led to a maximum individual population that occurred during peak species richness. There appears to be an optimum body size for each order sampled. While this information is interesting and might explain the abundance of hexapods, it does not address the factors that actually limit size.
Nanosella photo copyright © 1997 W. Eugene Hall
Page copyright 1998, David Bellamy.{kind=link}